

Sakshi Education
It is predominantly seen in prokaryotes where it accounts to 1% of the genome. Modified versions of the signaling system are also seen in fungi and plants. A pair of highly conserved proteins termed two-component regulators are employed which include, a sensor or histidine kinase (HK), being the primary component and an effector or response regulator (RR) as the secondary component. Two-component regulatory systems are defined by the presence of conserved blocks of sequence within the HK and RR, which are required for the signal transduction mechanisms.
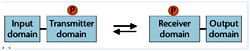
The HK is usually a membrane-spanning protein consisting of two domains- a N terminal sensory or input domain and a C terminal catalytic transmitter domain linked to a by a flexible linker. Most often, the sensory domain is extracytoplasmic by virtue of two transmembrane domains where it is ideally situated to sense a specific environmental parameter and communicates this information to the cytoplasmic RR protein through phospho transfer reactions.
Some HKs contain multiple membrane-spanning sequences which appear to be important for sensing signals localized to the membrane. Remarkably, some HKs are soluble proteins with N-terminal sensory domains that are involved in sensing intracellular signals. The N-terminal sensory domains exhibits a degree a variation and minimal or no homology among themselves, this variation enables them detect and interact with a wide variety of stimuli.
On the contrary, the C-terminal or kinase domains of most HKs exhibit a homology over a stretch of approximately350 amino acids. This corresponds to an independently folding domain responsible for Mg2+ and ATP binding and the catalysis of autophosphorylation. Although the overall level of sequence identity among kinase domains is low, there are several conserved boxes of amino acids that contain a few residues that are invariant in almost all members. RRs are generally soluble proteins whose activity is altered upon phosphorylation. They consist of N-terminal receiver domains responsible for mediating the phosphotransfer reactions with the cognate HK and C-terminal effector or output domains joined by a flexible linker region.
Unlike sensory domains, the RR’s show a certain homology over a particular region in N terminal receiver domains as well as C terminal region, which is perceived as requirement for DNA bonding? The first component acting as sensor perceives an input signal in the form of binding to a ligand or signal molecule. Oxygen availability, osmolarity, pH, ion levels and presence of pheromones are some examples of signals detected by the sensory domain. Upon ligand binding, the sensory domain gets activated by movement of one monomer relative to the other within a dimer exhibiting intrinsic kinase activity causing auto phosphorylation of the conserved histidine in the transmitter domain. The signal is subsequently relayed through phosphotransfer to a conserved aspartate residue in the receiver domain of the response regulator.
The intrinsic phosphotransfer enzyme activity is bidirectional on these proteins and several can be coupled in a chain. Thus, adaptive changes in the structure, behavior and physiology of the bacterial cell in response to a given cue can be initiated through signal induced alterations in a simple, conserved set of phospho transfer reactions between the HK and RR.
Mechanism of Signal Transduction
 Catlett et al. (2003), Eukaryotic Cell, vol 2(6), 1151–1161.
Catlett et al. (2003), Eukaryotic Cell, vol 2(6), 1151–1161.
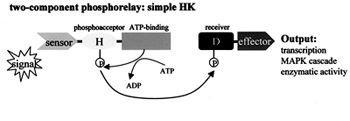
First, the HK functions as an auto kinase utilizing adenosine triphosphate (ATP) as the phosphate donor to phosphorylate itself on a conserved histidine (His) residue. The auto phosphorylation reaction occurs in trans between two subunits,of a dimer. The phosphohistidine bond has a high free energy and in the presence of the cognate RR, the phosphate is readily transferred to the conserved aspartate (Asp) residue within this protein. RR's can also exhibit kinase activity independently even in the absence of cognate HKs. They are capable of utilizing small molecules such as acetyl phosphate as phosphate donors. Thus they play a key role in phospho relay mechanism.
The degree of phosphorylation resulting in the formation of phosphorylated RR (RR–P) is kept under check through dephosphorylation events. Phosphatase activity is required for dephosphorylation that may be inherently present in the RRs themselves or HKs binding to the cognate RR-P might perform the activity. Upon phosphorylation conformational changes are induced in the RR's that lead to its activation and a concomitant response by affecting transcription. They interact with RNA polymerase with their phopsphorylated C-terminal or output domains.
RRs are grouped into three classes- OmpR, NtrC and FixJ based on the sequence similarity in the output domains. OmpR and FixJ class of RRs activate transcription by interacting with sigma-70 as a part of RNA polymerase holoenzyme. Whereas the Ntr C class of RRs interact with sigma 54 as a part of RNA polymerase holoenzyme.
Two Component System based Signaling in Plants
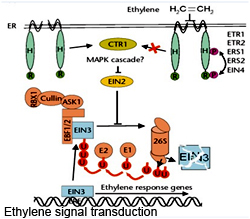 In Arabidopsis, eight two-component input regulator genes have been functionally identified, five for ethylene hormone perception and three for cytokinin. The ethylene receptors: ETR1, ETR2, ERS1, ERS2 and EIN4 share similarities with the histidine kinases. These receptors exhibit a positive regulation but in a negative manner. Gene transcription occurs only in the presence of the chemical signal and transcription is shut down in the absence of the signal. Since the signal/ ligand, Ethylene, being a gaseous hormone, the receptors are localized intracellularly on the surface of endoplasmic reticulum. These receptors are active constitutively and stimulate a protein kinase CONSTITUTIVE ETHYLENE RESPONSE1 (CTR1).
In Arabidopsis, eight two-component input regulator genes have been functionally identified, five for ethylene hormone perception and three for cytokinin. The ethylene receptors: ETR1, ETR2, ERS1, ERS2 and EIN4 share similarities with the histidine kinases. These receptors exhibit a positive regulation but in a negative manner. Gene transcription occurs only in the presence of the chemical signal and transcription is shut down in the absence of the signal. Since the signal/ ligand, Ethylene, being a gaseous hormone, the receptors are localized intracellularly on the surface of endoplasmic reticulum. These receptors are active constitutively and stimulate a protein kinase CONSTITUTIVE ETHYLENE RESPONSE1 (CTR1).
The exact mechanism of stimulation is not clear. Moreover these proteins are similar to the animal RAF1, a MAPK kinase kinase. CTR1 by an unknown mechanism blocks Elevator-InterNeuron (EIN2) another ER-located protein. EIN2 inhibits the 26S proteosome-mediated degradation of the transcription factor, EIN3, which activates the expression of genes in response to ethylene binding. EIN3 proteolysis is regulated by its association with other proteins like EBF1/2, F-box proteins present in the SCF-type E3 ubiquitin ligase complex.Therefore in the absence of ethylene, inactive EIN2 and active proteosome cause degradation of EIN3and thereby prevent transcription. Binding of Ethylene to the receptors inhibits the stimulation of CTR1, leading to activation of EIN2 and subsequently inhibits proteosomal degradation of EIN3.This facilitates accumulation of EIN3 to sufficient levels allowing transcription of ethylene response genes, assisted by EIN3.
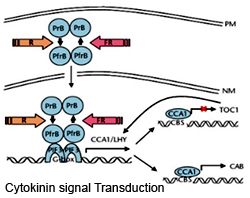 The cytokinin receptors also belong to the two-component regulators that bind cytokinin at their extracellular input domain .Binding of cytokinins results in a phospho-relay between the transmitter(H) and receiver(R)domains leading to the auto phosphorylation and activation of the receptor (see figure below). The activated receptor further stimulates the Arabidopsis histidine phosphotransfer protein (AHP) by phosphorylation which then translocated in the nucleus surpassing the nuclear membrane. In the nucleus it specifically interacts with the type-B Arabidopsis response regulators (ARRs) which bind to the DNA to activate the expression of cytokinin-response genes.
The cytokinin receptors also belong to the two-component regulators that bind cytokinin at their extracellular input domain .Binding of cytokinins results in a phospho-relay between the transmitter(H) and receiver(R)domains leading to the auto phosphorylation and activation of the receptor (see figure below). The activated receptor further stimulates the Arabidopsis histidine phosphotransfer protein (AHP) by phosphorylation which then translocated in the nucleus surpassing the nuclear membrane. In the nucleus it specifically interacts with the type-B Arabidopsis response regulators (ARRs) which bind to the DNA to activate the expression of cytokinin-response genes.
References:
The HK is usually a membrane-spanning protein consisting of two domains- a N terminal sensory or input domain and a C terminal catalytic transmitter domain linked to a by a flexible linker. Most often, the sensory domain is extracytoplasmic by virtue of two transmembrane domains where it is ideally situated to sense a specific environmental parameter and communicates this information to the cytoplasmic RR protein through phospho transfer reactions.
Some HKs contain multiple membrane-spanning sequences which appear to be important for sensing signals localized to the membrane. Remarkably, some HKs are soluble proteins with N-terminal sensory domains that are involved in sensing intracellular signals. The N-terminal sensory domains exhibits a degree a variation and minimal or no homology among themselves, this variation enables them detect and interact with a wide variety of stimuli.
On the contrary, the C-terminal or kinase domains of most HKs exhibit a homology over a stretch of approximately350 amino acids. This corresponds to an independently folding domain responsible for Mg2+ and ATP binding and the catalysis of autophosphorylation. Although the overall level of sequence identity among kinase domains is low, there are several conserved boxes of amino acids that contain a few residues that are invariant in almost all members. RRs are generally soluble proteins whose activity is altered upon phosphorylation. They consist of N-terminal receiver domains responsible for mediating the phosphotransfer reactions with the cognate HK and C-terminal effector or output domains joined by a flexible linker region.
Unlike sensory domains, the RR’s show a certain homology over a particular region in N terminal receiver domains as well as C terminal region, which is perceived as requirement for DNA bonding? The first component acting as sensor perceives an input signal in the form of binding to a ligand or signal molecule. Oxygen availability, osmolarity, pH, ion levels and presence of pheromones are some examples of signals detected by the sensory domain. Upon ligand binding, the sensory domain gets activated by movement of one monomer relative to the other within a dimer exhibiting intrinsic kinase activity causing auto phosphorylation of the conserved histidine in the transmitter domain. The signal is subsequently relayed through phosphotransfer to a conserved aspartate residue in the receiver domain of the response regulator.
The intrinsic phosphotransfer enzyme activity is bidirectional on these proteins and several can be coupled in a chain. Thus, adaptive changes in the structure, behavior and physiology of the bacterial cell in response to a given cue can be initiated through signal induced alterations in a simple, conserved set of phospho transfer reactions between the HK and RR.
Mechanism of Signal Transduction
First, the HK functions as an auto kinase utilizing adenosine triphosphate (ATP) as the phosphate donor to phosphorylate itself on a conserved histidine (His) residue. The auto phosphorylation reaction occurs in trans between two subunits,of a dimer. The phosphohistidine bond has a high free energy and in the presence of the cognate RR, the phosphate is readily transferred to the conserved aspartate (Asp) residue within this protein. RR's can also exhibit kinase activity independently even in the absence of cognate HKs. They are capable of utilizing small molecules such as acetyl phosphate as phosphate donors. Thus they play a key role in phospho relay mechanism.
The degree of phosphorylation resulting in the formation of phosphorylated RR (RR–P) is kept under check through dephosphorylation events. Phosphatase activity is required for dephosphorylation that may be inherently present in the RRs themselves or HKs binding to the cognate RR-P might perform the activity. Upon phosphorylation conformational changes are induced in the RR's that lead to its activation and a concomitant response by affecting transcription. They interact with RNA polymerase with their phopsphorylated C-terminal or output domains.
RRs are grouped into three classes- OmpR, NtrC and FixJ based on the sequence similarity in the output domains. OmpR and FixJ class of RRs activate transcription by interacting with sigma-70 as a part of RNA polymerase holoenzyme. Whereas the Ntr C class of RRs interact with sigma 54 as a part of RNA polymerase holoenzyme.
Two Component System based Signaling in Plants
In Arabidopsis, eight two-component input regulator genes have been functionally identified, five for ethylene hormone perception and three for cytokinin. The ethylene receptors: ETR1, ETR2, ERS1, ERS2 and EIN4 share similarities with the histidine kinases. These receptors exhibit a positive regulation but in a negative manner. Gene transcription occurs only in the presence of the chemical signal and transcription is shut down in the absence of the signal. Since the signal/ ligand, Ethylene, being a gaseous hormone, the receptors are localized intracellularly on the surface of endoplasmic reticulum. These receptors are active constitutively and stimulate a protein kinase CONSTITUTIVE ETHYLENE RESPONSE1 (CTR1). The exact mechanism of stimulation is not clear. Moreover these proteins are similar to the animal RAF1, a MAPK kinase kinase. CTR1 by an unknown mechanism blocks Elevator-InterNeuron (EIN2) another ER-located protein. EIN2 inhibits the 26S proteosome-mediated degradation of the transcription factor, EIN3, which activates the expression of genes in response to ethylene binding. EIN3 proteolysis is regulated by its association with other proteins like EBF1/2, F-box proteins present in the SCF-type E3 ubiquitin ligase complex.Therefore in the absence of ethylene, inactive EIN2 and active proteosome cause degradation of EIN3and thereby prevent transcription. Binding of Ethylene to the receptors inhibits the stimulation of CTR1, leading to activation of EIN2 and subsequently inhibits proteosomal degradation of EIN3.This facilitates accumulation of EIN3 to sufficient levels allowing transcription of ethylene response genes, assisted by EIN3.
The cytokinin receptors also belong to the two-component regulators that bind cytokinin at their extracellular input domain .Binding of cytokinins results in a phospho-relay between the transmitter(H) and receiver(R)domains leading to the auto phosphorylation and activation of the receptor (see figure below). The activated receptor further stimulates the Arabidopsis histidine phosphotransfer protein (AHP) by phosphorylation which then translocated in the nucleus surpassing the nuclear membrane. In the nucleus it specifically interacts with the type-B Arabidopsis response regulators (ARRs) which bind to the DNA to activate the expression of cytokinin-response genes. References:
- Helariutta .Y(2006) Signs of Change: Hormone Receptors that Regulate the Plant Development. Development 133; 1857-1869.
- Johnson KL and Ingram GC (2005) Sending the Right Signals: Regulating Receptor Kinase Activity. Current Opinion in Plant Biology 8; 648-656.
- Bogre L et al. (2004) Learning the Lipid Language of Plant Signaling. Trends in Plant Science 9; 378-384.
- Henriques R et al. (2003) Growth Signaling Pathways in Arabidopsis and the AGC Protein Kinases. Trends in Plant Science 8; 424-431.
- Catlett et al. (2003), Whole-Genome Analysis of Two-Component Signal Transduction Genes in Fungal Pathogens. Eukaryotic Cell, 2(6), 1151–1161.
- Hwang I, Chen HC and Sheen J (2002) Two Component Signal Transduction Pathways in Arabidopsis. Plant Physiology 129; 500-515.
Published date : 14 May 2014 02:37PM




More Articles


