

Sakshi Education
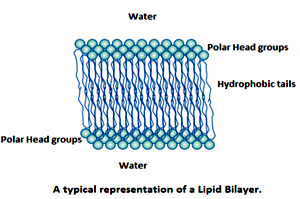
A plasma membrane is a thin barrier that separates the interior contents of the cell from the exterior. It would be requiring about five thousand plasma membranes stacked one on top of the other to equal the thickness of a single page of the book. In eukaryotic cells it also partitions the cytoplasm into internal compartments called organelles like nucleus, mitochondria, chloroplasts etc.
Biological membranes act as selective permeability barriers that allow the directional transport of some types of molecules but exclude the transport of others. Two major components constitute the cell membrane structure- lipid bilayer and membrane proteins. They execute majority of the membrane functions apart from acting as permeability barriers controlling the transport of molecules into and out of the cell, transmitting signals from the environment to the cell interior, participate in the synthesis and assembly of cell wall molecules, and provides physical link between elements of cytoskeleton and the Extra Cellular Matrix. The plasma membrane produces Plasmodesmata, membrane tubes that cross wall and provides direct channels of communication between adjacent cells. Where virtually every cell has separate plasma membrane and cell to cell communication occur instead through protein channels known as gap junctions. Animal cells are normally under isosmotic pressure with their environment but plant cells experience a turgor pressure, where the plasma membrane is held tightly against the cell wall. Lipids present in the membrane also act as a source of ‘lipid second messengers’ such as diacylglycerol, lysophosphatidicacid, eicosanoids, inositol trisphosphate, ceramide and sphingosine.
Salient features of biological membranes-
Membrane lipids contain a hydrophobic (water-hating) portion and a hydrophilic (water-loving) component and are a thus amphipathic in nature. The hydrophobic domains of lipids spontaneously segregate away from the aqueous compartment, whereas the hydrophilic head groups are oriented away from the hydrophobic environment and are in contact with the aqueous milieu. Thus, amphipathic lipids have a tendency to self-associate non-covalently in an aqueous environment such as that encountered in a cell to form a lipid bilayer. In simple terms the amphipathic membrane lipids in an aqueous environment spontaneously form a bilayer structure. Hence the structure of a lipid bilayer is driven by the chemical structure of the lipid molecules. Thus the bilayer structure constitutes the basic structure of all cell membranes and viral envelopes.
If isolated biological lipids or pure synthetic lipids are placed in water, liposomes, containing concentric layers of lipid bilayer are formed. Amphipathic lipid molecules including phospholipids and cholesterol, apart from forming lipid bilayers, can also form lipid monolayers on the surface of serum lipoproteins. Another biologically important structure is the micelle Detergents form micelles. Molecules that contain a larger polar head than the hydrophobic tails form micelles like a cone. Packing if cones with the tails embedded inside to form a sphere results in a micelle formation.
A biological example of micelles is found in the digestive tract where micelles containing bile salts, solubilize fat (triglycerides) in the gut from the diet
The lipid composition of plasma membranes is highly variable. There are three major types of lipids in plasma membranes, namely Phospholipids, Spingolipids and Sterol.
Phospholipids:
The most abundant and major class of membrane lipids. These have a polar head group and two hydrophobic hydrocarbon tails. The tails usually fatty acid chain and they differ in length (they normally contain between14 and 24carbon atoms) and c-16 and c-18 carbon contain fatty acids are common. The alkyl chain may be saturated (or) may contain more double bonds. The confirmation of the double bonds is most unsaturated fatty acid is Cis. The double bonds in poly unsaturated fatty acids are separated by at least one markedly dependent on chain length and degree of unsaturation. Unsaturated fatty acids have lower melting point than saturated fatty acids of the same length.
A phospholipid molecule constructed from four components. Those are one or more fatty acid chain, phosphate, alcohol and a plat form to which the fatty acids are attached (Glycerol).
The fatty acid components provide a hydrophobic barrier, whereas the remainder of the molecule has hydrophilic properties to enable interaction with the aqueous environment.
Phospholipids derived from glycerol are called phoshpogycerides.
In this the hydroxyl group of (-OH) at C-1 and C-2 of glycerol are esterified to the carboxyl group of the two fatty acids chains. The C-3 hydroxyl group of the glycerol backbone is esterified to phosphoric acid. When no further additions are made, the resulting compound is Phoshotidate (Di Acyl Glycerol) the simplest phoshpogycerides. Only small amount of phoshotidate are present in membranes, and it act as a key intermediate in the biosynthesis of the other phoshpogycerides.
Shape and amphipathic nature of the lipid molecule that causes to form bilayer spontaneously in aqueous solution. Depending shape of the lipid molecule forms to structure, Micelles (with tails inwards), bimolecular sheets and bilayers. Cylindrical shape phospholipid molecules spontaneously form bilayer in aqueous environment. Moreover, these lipid bilayers tend to close on themselves to form sealed compartments, thereby eliminating free edges where the hydrophobic tail would in contact with water.
A lipid bilayer has self- sealing properties that make an ideal structure for cell membranes. Two types of synthetic bilayers have been very useful in experimental studies. Those are Liposomes (spherical vesicles 25nm to 1µm in diameter) and Black membrane (planar bilayer, formed across a hole in a partition between two aqueous compartments and used to measure the permeability properties of synthetic membranes.
The alcohol head groups rather than fatty acids, gives their phoshpogycerides their names:
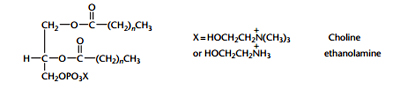 All of the Phospholipids have negative charges on the phosphate esterified to the alcohols. And neutral phoshpogycerides are Phosphotidyl choline and Phosphotidyl ethanolamine have a positive charge on their nitrogen giving them a net charge of zero (0).
All of the Phospholipids have negative charges on the phosphate esterified to the alcohols. And neutral phoshpogycerides are Phosphotidyl choline and Phosphotidyl ethanolamine have a positive charge on their nitrogen giving them a net charge of zero (0).
Phosphotidyl Serine has extra positive and negative charges, giving it a net negative like other acidic phoshpogycerides, due to presence of negative charge; it always remains in inner leaflet of plasma membrane but moved outer layer during apoptosis, it may serves to attached phagocytes to consume cell remnants after the apoptosis is completed. This can be synthesized by Base Exchange reaction.
A phospholipids molecule in synthetic bilayer very rarely migrates from the monolayer on one side to that on the outer side. Flip flop movement- Occur less than once in month for any individual molecules. Lipid molecules readily exchanges place their neighbour with in a monolayer called rapid lateral diffusion. Individual lipid molecules rotate very rapidly about their long axis and that their hydrocarbon chain flexible.
This lipid component of biological membrane is a two dimensional liquid in which the construing molecules are free move laterally. Phospholipid molecules are synthesized in only in one monolayer of membrane, mainly being produced in the cytosolic monolayer of the Endoplasmic Reticulum, newly made molecules could migrates to that other half of the lipid bilayer.ER membrane bound enzymes called Phospholipid trans locators, which catalysed the rapid flip flop of specific phospholipid from the monolayer where they are made to opposite monolayer.
Spingolipids:
The sphingolipids constitute a class of lipids that are components of membranes of animals, plants, fungi and a few prokaryotes. Most sugars containing lipids of biological membranes are Spingolipids. They are the structural counterpart of glycerol and one fatty acid of phoshphogycerides. In mammals, these lipids are especially abundant in the brain and the nervous system. Sphingolipids are oriented asymmetrically in the membrane bilayer and are located predominantly on the outer leaflet of the plasma membrane with the polar head group facing towards the outside of the cell. However, some sphingolipids, such as ceramide and sphingosine, are also present in small amounts on intracellular membranes where they have been reported to mediate signal transduction events that regulate cell growth and survival.
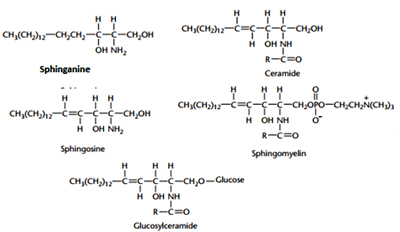 The basic building block of all sphingolipid molecules is a sphingoid base such as sphingosine or sphinganine. Sphingomyelin is a phosphosphingolipid that is particularly abundant in the myelin sheath that insulates nerve fibres. This lipid is synthesised by the transfer of phosphorylcholine from phosphatidylcholine to the terminal C-1 hydroxyl of ceramide in a reaction catalysed by sphingomyelin synthase. The structures of the glycosphingolipids are remarkably diverse but all are built upon ceramide. For example, the hydroxyl group at carbon-1 of ceramide can be linked in a glycosidic linkage either to a single carbohydrate molecule (glucose in glucosylceramide or galactose in galactosylceramide) or to an oligosaccharide chain. The most common carbohydrates that occur in the glycosphingolipids are glucose, galactose, N-acetylglucosamine, N-acetylgalactosamine, fucose and mannose. Some of the fucose-containing glycosphingolipids are blood group antigens. The gangliosides comprise a subclass of the glycosphingolipids that are defined by the presence of one or more sialic acid molecules in the oliogosaccharide portion of the molecule. A defective breakdown of individual glycosphingolipid species due to lack of sphingolipidoses, cause a group of human diseases like Tay sach’s, Niemann Pick A Disease and Fabry’s disease.
The basic building block of all sphingolipid molecules is a sphingoid base such as sphingosine or sphinganine. Sphingomyelin is a phosphosphingolipid that is particularly abundant in the myelin sheath that insulates nerve fibres. This lipid is synthesised by the transfer of phosphorylcholine from phosphatidylcholine to the terminal C-1 hydroxyl of ceramide in a reaction catalysed by sphingomyelin synthase. The structures of the glycosphingolipids are remarkably diverse but all are built upon ceramide. For example, the hydroxyl group at carbon-1 of ceramide can be linked in a glycosidic linkage either to a single carbohydrate molecule (glucose in glucosylceramide or galactose in galactosylceramide) or to an oligosaccharide chain. The most common carbohydrates that occur in the glycosphingolipids are glucose, galactose, N-acetylglucosamine, N-acetylgalactosamine, fucose and mannose. Some of the fucose-containing glycosphingolipids are blood group antigens. The gangliosides comprise a subclass of the glycosphingolipids that are defined by the presence of one or more sialic acid molecules in the oliogosaccharide portion of the molecule. A defective breakdown of individual glycosphingolipid species due to lack of sphingolipidoses, cause a group of human diseases like Tay sach’s, Niemann Pick A Disease and Fabry’s disease.
Tay-Sach’s disease occurs when harmful quantities of gangliosides accumulate in the brain's nerve cells, eventually leading to the premature death of the cells. Niemann Picks Disease refers to a group of inherited severe metabolic disorders that allow sphingomyelin to accumulate in lysosomes, which are organelles in animal cells. The severe form is fatal in toddlerhood; people with milder forms may live into their teens or young adulthood. It is a kind of sphingolipidosis. Sphingolipidoses, in turn, are included in the larger family of lysosomal storage diseases (LSDs). Fabry’s diseaseis a rare genetic lysosomal storage disease, inherited in an X-linked manner. Fabry disease can cause a wide range of systemic symptoms.
Sterols:
Sterols are third major class of membrane lipids. Cholesterol is the major steroid in animal plasma membranes, with lower concentration of in internal membranes. Plants, lower eukaryotes, and bacteria have other sterols in their membranes, which are Stigmosterols, sistosterols and Champ sterols. Cholesterol is a polar, so it is inserts into the core of bilayers with the hydroxyl on C3 oriented towards the face.
Archaeal membranes are built from Ether lipids with branched chains of fatty acids. The nonpolar chains are joined to a glycerol backbone by ether rather than ester linkage. The ether linkage more resistant to hydrolysis. The alkyl chains are branched rather than linear. They are built up from repeats of a fully saturated five carbon fragment. These branched saturated hydrocarbons are more resistant to oxidation. The ability of Archaeal lipid to resist hydrolysis and oxidation may help these organisms to withstand the extreme conditions, such as high temperature, low pH or high salt concentration, under which some of these Archaeal grow.
Fluidity of lipid bilayer:
The fluidity of plasma membrane is depends on lipid composition temperature and saturation, length of fatty acid chain. In general, an increase in the temperature tends to elevate membrane fluidity. Lowering of temperature causes a freely flowing, low viscosity, fluid like membrane structure to a more rigid, gel like organisation. Lipid with short or unsaturated fatty acyl chains undergo the phase transition at lower temperatures than lipid with long or saturated chains.
Shorter chain length reduces the tendency of the hydrocarbon tails to interact with one another. Unsaturated fatty acyl chain have kink, thus tend to adopt to increase fluidity of plasma membranes.
Cholesterol is also major detriment of membrane fluidity. At high temperature, cholesterol interfaces with the movement of the phospholipid fatty acid chains, making the less fluid but at low temperature in interfaces interaction between fatty acid chains and prevents membranes from freezing. At high concentration found in eukaryotic plasma membranes, cholesterol makes to membrane over all less fluid at growth temperatures near 37°C.
Membranes act as ordered fluids:
The lipid bilayer of cellular membranes allows lateral movement, or diffusion of lipids in the plane of the membrane but movement in and out of the plane of the membrane is limited by the hydrophobic effect, establishing a degree if orderliness in the structure. Also side-by-side arrangement of the lipids in the bilayer also imposes another degree of order. There exists a greater conformational flexibility of the hydrocarbon chain in the centre of the bilayer than in the region near the lipid head groups due to the intramolecular interactions. Lateral diffusion of the proteins also occurs to enable some membrane functions.
Membranes exhibit Limited permeability:
Membranes are impermeable to solutes and polar molecules. Therefore transport of such molecules is mediated by membrane proteins.
Membrane Proteins
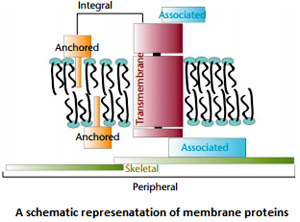 Membrane proteins can be classified as integral membrane proteins and peripheral membrane proteins. These classes of membrane proteins can be subdivided further as Anchored membrane proteins insert a hydrophobic portion into one leaflet of the bilayer and include proteins that have hydrophobic lipid covalently attached. Associated proteins are peripheral membrane proteins that are bound to integral membrane proteins, and skeletal membrane proteins form a network underlying the plasma membrane of a cell.
Membrane proteins can be classified as integral membrane proteins and peripheral membrane proteins. These classes of membrane proteins can be subdivided further as Anchored membrane proteins insert a hydrophobic portion into one leaflet of the bilayer and include proteins that have hydrophobic lipid covalently attached. Associated proteins are peripheral membrane proteins that are bound to integral membrane proteins, and skeletal membrane proteins form a network underlying the plasma membrane of a cell.
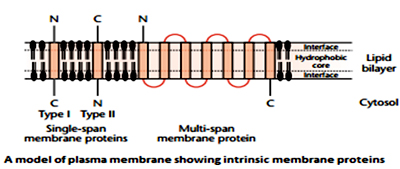 Integral membrane proteins can be further classified as transmembrane proteins or anchored proteins. Transmembrane proteins completely traverse the lipid bilayer and expose some of their mass on both sides of the membrane. An example of such a protein is the G protein coupled receptor that transduces signals from one side of the membrane to the other.
Integral membrane proteins can be further classified as transmembrane proteins or anchored proteins. Transmembrane proteins completely traverse the lipid bilayer and expose some of their mass on both sides of the membrane. An example of such a protein is the G protein coupled receptor that transduces signals from one side of the membrane to the other.
Anchored membrane proteins have part of their mass buried within the hydrophobic part of the lipid bilayer, but with structures that do not completely traverse the membrane. An example can be found in the lipid anchored proteins likeThy1that resemble soluble proteins but are covalently linked to an amphipathic membrane lipid that anchors the protein into the outer leaflet of the plasma membrane.
Peripheral membrane proteins are associated with cell membranes, but do not significantly penetrate the hydrophobic interior of the lipid bilayer. They generally resemble water-soluble proteins in structure. Peripheral membrane proteins can be classified as associated membrane proteins or as membrane skeleton. Associated membrane proteins bind to integral membrane proteins (or to the surface of the lipid bilayer through electrostatic interactions with lipid head groups).They may form part of a functioning complex with the integral membrane protein. An example is cytochrome c, which binds to cytochrome c oxidase (an integral membrane protein in the inner mitochondrial membrane) to donate an electron as part of the electron transport chain supporting the synthesis of ATP in the inner mitochondrial membrane. Proteins can also form a skeleton lining the inside of the plasma membrane of cells by binding either to integral membrane proteins or associated proteins.
Spectrin present in the inner side (cytoplasmic) of the plasma membrane of RBCs binds to ankyrin, an associated protein, which in turn binds to band 3, a transmembrane protein. Skeletal proteins can restrict the lateral diffusion of integral proteins.
References:
Biological membranes act as selective permeability barriers that allow the directional transport of some types of molecules but exclude the transport of others. Two major components constitute the cell membrane structure- lipid bilayer and membrane proteins. They execute majority of the membrane functions apart from acting as permeability barriers controlling the transport of molecules into and out of the cell, transmitting signals from the environment to the cell interior, participate in the synthesis and assembly of cell wall molecules, and provides physical link between elements of cytoskeleton and the Extra Cellular Matrix. The plasma membrane produces Plasmodesmata, membrane tubes that cross wall and provides direct channels of communication between adjacent cells. Where virtually every cell has separate plasma membrane and cell to cell communication occur instead through protein channels known as gap junctions. Animal cells are normally under isosmotic pressure with their environment but plant cells experience a turgor pressure, where the plasma membrane is held tightly against the cell wall. Lipids present in the membrane also act as a source of ‘lipid second messengers’ such as diacylglycerol, lysophosphatidicacid, eicosanoids, inositol trisphosphate, ceramide and sphingosine.
Salient features of biological membranes-
- Membranes are sheet like structures, only two molecules thick, that form closed boundaries between different compartments. The thickness of most membranes is between 6nm and 10nm.
- Membranes consist mainly of lipids and proteins. The mass ratio of lipid to protein ranges from 1:4 to 4:1. Membranes also contain carbohydrates that are linked to lipids and proteins.
- Membranes lipids are small molecules that have both hydrophilic and hydrophobic moieties. These lipids spontaneously form closed bimolecular sheets in aqueous media. These lipid bilayers are barriers to the flow of polar molecules.
- Membrane proteins are embedded in lipid bilayers, which create suitable environments for their action like acts as pumps, channels, carrier, receptor, energy transducers and enzymes.
- Membranes are non- covalent assemblies, Membranes are asymmetric in the distribution of proteins and thus the faces of biological membranes always differ from each other.
- Membranes can be regarded as two dimensional solutions of oriented proteins and lipids.
- Most cell membranes are electrically polarized, such that the inside is negative [typically -60 mill Volts (mV)]; membrane potential is play key role in transport, energy conversion and excitability.
- Daughter cells inherited a complete set of membranes from their mother.
- Each potential mother cell maintains a complete set of membranes.
- New membranes can be produced only by growth and fission of existing membranes, they cannot arise de novo.
Membrane lipids contain a hydrophobic (water-hating) portion and a hydrophilic (water-loving) component and are a thus amphipathic in nature. The hydrophobic domains of lipids spontaneously segregate away from the aqueous compartment, whereas the hydrophilic head groups are oriented away from the hydrophobic environment and are in contact with the aqueous milieu. Thus, amphipathic lipids have a tendency to self-associate non-covalently in an aqueous environment such as that encountered in a cell to form a lipid bilayer. In simple terms the amphipathic membrane lipids in an aqueous environment spontaneously form a bilayer structure. Hence the structure of a lipid bilayer is driven by the chemical structure of the lipid molecules. Thus the bilayer structure constitutes the basic structure of all cell membranes and viral envelopes.
If isolated biological lipids or pure synthetic lipids are placed in water, liposomes, containing concentric layers of lipid bilayer are formed. Amphipathic lipid molecules including phospholipids and cholesterol, apart from forming lipid bilayers, can also form lipid monolayers on the surface of serum lipoproteins. Another biologically important structure is the micelle Detergents form micelles. Molecules that contain a larger polar head than the hydrophobic tails form micelles like a cone. Packing if cones with the tails embedded inside to form a sphere results in a micelle formation.
A biological example of micelles is found in the digestive tract where micelles containing bile salts, solubilize fat (triglycerides) in the gut from the diet
The lipid composition of plasma membranes is highly variable. There are three major types of lipids in plasma membranes, namely Phospholipids, Spingolipids and Sterol.
Phospholipids:
The most abundant and major class of membrane lipids. These have a polar head group and two hydrophobic hydrocarbon tails. The tails usually fatty acid chain and they differ in length (they normally contain between14 and 24carbon atoms) and c-16 and c-18 carbon contain fatty acids are common. The alkyl chain may be saturated (or) may contain more double bonds. The confirmation of the double bonds is most unsaturated fatty acid is Cis. The double bonds in poly unsaturated fatty acids are separated by at least one markedly dependent on chain length and degree of unsaturation. Unsaturated fatty acids have lower melting point than saturated fatty acids of the same length.
A phospholipid molecule constructed from four components. Those are one or more fatty acid chain, phosphate, alcohol and a plat form to which the fatty acids are attached (Glycerol).
The fatty acid components provide a hydrophobic barrier, whereas the remainder of the molecule has hydrophilic properties to enable interaction with the aqueous environment.
Phospholipids derived from glycerol are called phoshpogycerides.
In this the hydroxyl group of (-OH) at C-1 and C-2 of glycerol are esterified to the carboxyl group of the two fatty acids chains. The C-3 hydroxyl group of the glycerol backbone is esterified to phosphoric acid. When no further additions are made, the resulting compound is Phoshotidate (Di Acyl Glycerol) the simplest phoshpogycerides. Only small amount of phoshotidate are present in membranes, and it act as a key intermediate in the biosynthesis of the other phoshpogycerides.
Shape and amphipathic nature of the lipid molecule that causes to form bilayer spontaneously in aqueous solution. Depending shape of the lipid molecule forms to structure, Micelles (with tails inwards), bimolecular sheets and bilayers. Cylindrical shape phospholipid molecules spontaneously form bilayer in aqueous environment. Moreover, these lipid bilayers tend to close on themselves to form sealed compartments, thereby eliminating free edges where the hydrophobic tail would in contact with water.
A lipid bilayer has self- sealing properties that make an ideal structure for cell membranes. Two types of synthetic bilayers have been very useful in experimental studies. Those are Liposomes (spherical vesicles 25nm to 1µm in diameter) and Black membrane (planar bilayer, formed across a hole in a partition between two aqueous compartments and used to measure the permeability properties of synthetic membranes.
The alcohol head groups rather than fatty acids, gives their phoshpogycerides their names:
- Phosphatidic Acid (PA)- no head group.
- Phosphotidyl Glycerol(PG)-glycerol head group
- Phosphotidyl Ethanolamine (PE)- Ethanolamine head group, abundant in nerve tissues
- Phosphotidyl Choline (PC)- choline head group, abundant in heart tissues.
- Phosphotidyl Serine (PS)- serine head group
- Phosphotidyl Inositol (PI). Inositol head group.
All of the Phospholipids have negative charges on the phosphate esterified to the alcohols. And neutral phoshpogycerides are Phosphotidyl choline and Phosphotidyl ethanolamine have a positive charge on their nitrogen giving them a net charge of zero (0).Phosphotidyl Serine has extra positive and negative charges, giving it a net negative like other acidic phoshpogycerides, due to presence of negative charge; it always remains in inner leaflet of plasma membrane but moved outer layer during apoptosis, it may serves to attached phagocytes to consume cell remnants after the apoptosis is completed. This can be synthesized by Base Exchange reaction.
A phospholipids molecule in synthetic bilayer very rarely migrates from the monolayer on one side to that on the outer side. Flip flop movement- Occur less than once in month for any individual molecules. Lipid molecules readily exchanges place their neighbour with in a monolayer called rapid lateral diffusion. Individual lipid molecules rotate very rapidly about their long axis and that their hydrocarbon chain flexible.
This lipid component of biological membrane is a two dimensional liquid in which the construing molecules are free move laterally. Phospholipid molecules are synthesized in only in one monolayer of membrane, mainly being produced in the cytosolic monolayer of the Endoplasmic Reticulum, newly made molecules could migrates to that other half of the lipid bilayer.ER membrane bound enzymes called Phospholipid trans locators, which catalysed the rapid flip flop of specific phospholipid from the monolayer where they are made to opposite monolayer.
Spingolipids:
The sphingolipids constitute a class of lipids that are components of membranes of animals, plants, fungi and a few prokaryotes. Most sugars containing lipids of biological membranes are Spingolipids. They are the structural counterpart of glycerol and one fatty acid of phoshphogycerides. In mammals, these lipids are especially abundant in the brain and the nervous system. Sphingolipids are oriented asymmetrically in the membrane bilayer and are located predominantly on the outer leaflet of the plasma membrane with the polar head group facing towards the outside of the cell. However, some sphingolipids, such as ceramide and sphingosine, are also present in small amounts on intracellular membranes where they have been reported to mediate signal transduction events that regulate cell growth and survival.
The basic building block of all sphingolipid molecules is a sphingoid base such as sphingosine or sphinganine. Sphingomyelin is a phosphosphingolipid that is particularly abundant in the myelin sheath that insulates nerve fibres. This lipid is synthesised by the transfer of phosphorylcholine from phosphatidylcholine to the terminal C-1 hydroxyl of ceramide in a reaction catalysed by sphingomyelin synthase. The structures of the glycosphingolipids are remarkably diverse but all are built upon ceramide. For example, the hydroxyl group at carbon-1 of ceramide can be linked in a glycosidic linkage either to a single carbohydrate molecule (glucose in glucosylceramide or galactose in galactosylceramide) or to an oligosaccharide chain. The most common carbohydrates that occur in the glycosphingolipids are glucose, galactose, N-acetylglucosamine, N-acetylgalactosamine, fucose and mannose. Some of the fucose-containing glycosphingolipids are blood group antigens. The gangliosides comprise a subclass of the glycosphingolipids that are defined by the presence of one or more sialic acid molecules in the oliogosaccharide portion of the molecule. A defective breakdown of individual glycosphingolipid species due to lack of sphingolipidoses, cause a group of human diseases like Tay sach’s, Niemann Pick A Disease and Fabry’s disease. Tay-Sach’s disease occurs when harmful quantities of gangliosides accumulate in the brain's nerve cells, eventually leading to the premature death of the cells. Niemann Picks Disease refers to a group of inherited severe metabolic disorders that allow sphingomyelin to accumulate in lysosomes, which are organelles in animal cells. The severe form is fatal in toddlerhood; people with milder forms may live into their teens or young adulthood. It is a kind of sphingolipidosis. Sphingolipidoses, in turn, are included in the larger family of lysosomal storage diseases (LSDs). Fabry’s diseaseis a rare genetic lysosomal storage disease, inherited in an X-linked manner. Fabry disease can cause a wide range of systemic symptoms.
Sterols:
Sterols are third major class of membrane lipids. Cholesterol is the major steroid in animal plasma membranes, with lower concentration of in internal membranes. Plants, lower eukaryotes, and bacteria have other sterols in their membranes, which are Stigmosterols, sistosterols and Champ sterols. Cholesterol is a polar, so it is inserts into the core of bilayers with the hydroxyl on C3 oriented towards the face.
Archaeal membranes are built from Ether lipids with branched chains of fatty acids. The nonpolar chains are joined to a glycerol backbone by ether rather than ester linkage. The ether linkage more resistant to hydrolysis. The alkyl chains are branched rather than linear. They are built up from repeats of a fully saturated five carbon fragment. These branched saturated hydrocarbons are more resistant to oxidation. The ability of Archaeal lipid to resist hydrolysis and oxidation may help these organisms to withstand the extreme conditions, such as high temperature, low pH or high salt concentration, under which some of these Archaeal grow.
Fluidity of lipid bilayer:
The fluidity of plasma membrane is depends on lipid composition temperature and saturation, length of fatty acid chain. In general, an increase in the temperature tends to elevate membrane fluidity. Lowering of temperature causes a freely flowing, low viscosity, fluid like membrane structure to a more rigid, gel like organisation. Lipid with short or unsaturated fatty acyl chains undergo the phase transition at lower temperatures than lipid with long or saturated chains.
Shorter chain length reduces the tendency of the hydrocarbon tails to interact with one another. Unsaturated fatty acyl chain have kink, thus tend to adopt to increase fluidity of plasma membranes.
Cholesterol is also major detriment of membrane fluidity. At high temperature, cholesterol interfaces with the movement of the phospholipid fatty acid chains, making the less fluid but at low temperature in interfaces interaction between fatty acid chains and prevents membranes from freezing. At high concentration found in eukaryotic plasma membranes, cholesterol makes to membrane over all less fluid at growth temperatures near 37°C.
Membranes act as ordered fluids:
The lipid bilayer of cellular membranes allows lateral movement, or diffusion of lipids in the plane of the membrane but movement in and out of the plane of the membrane is limited by the hydrophobic effect, establishing a degree if orderliness in the structure. Also side-by-side arrangement of the lipids in the bilayer also imposes another degree of order. There exists a greater conformational flexibility of the hydrocarbon chain in the centre of the bilayer than in the region near the lipid head groups due to the intramolecular interactions. Lateral diffusion of the proteins also occurs to enable some membrane functions.
Membranes exhibit Limited permeability:
Membranes are impermeable to solutes and polar molecules. Therefore transport of such molecules is mediated by membrane proteins.
Membrane Proteins
Membrane proteins can be classified as integral membrane proteins and peripheral membrane proteins. These classes of membrane proteins can be subdivided further as Anchored membrane proteins insert a hydrophobic portion into one leaflet of the bilayer and include proteins that have hydrophobic lipid covalently attached. Associated proteins are peripheral membrane proteins that are bound to integral membrane proteins, and skeletal membrane proteins form a network underlying the plasma membrane of a cell. Integral membrane proteins can be further classified as transmembrane proteins or anchored proteins. Transmembrane proteins completely traverse the lipid bilayer and expose some of their mass on both sides of the membrane. An example of such a protein is the G protein coupled receptor that transduces signals from one side of the membrane to the other.Anchored membrane proteins have part of their mass buried within the hydrophobic part of the lipid bilayer, but with structures that do not completely traverse the membrane. An example can be found in the lipid anchored proteins likeThy1that resemble soluble proteins but are covalently linked to an amphipathic membrane lipid that anchors the protein into the outer leaflet of the plasma membrane.
Peripheral membrane proteins are associated with cell membranes, but do not significantly penetrate the hydrophobic interior of the lipid bilayer. They generally resemble water-soluble proteins in structure. Peripheral membrane proteins can be classified as associated membrane proteins or as membrane skeleton. Associated membrane proteins bind to integral membrane proteins (or to the surface of the lipid bilayer through electrostatic interactions with lipid head groups).They may form part of a functioning complex with the integral membrane protein. An example is cytochrome c, which binds to cytochrome c oxidase (an integral membrane protein in the inner mitochondrial membrane) to donate an electron as part of the electron transport chain supporting the synthesis of ATP in the inner mitochondrial membrane. Proteins can also form a skeleton lining the inside of the plasma membrane of cells by binding either to integral membrane proteins or associated proteins.
Spectrin present in the inner side (cytoplasmic) of the plasma membrane of RBCs binds to ankyrin, an associated protein, which in turn binds to band 3, a transmembrane protein. Skeletal proteins can restrict the lateral diffusion of integral proteins.
References:
- Alberts B, Johnson A, Lewis J et al. (2007) The Molecular Biology of the Cell, 5th edn. New York: Garland Press.
- Vance DE and Vance J (2008) Biochemistry of Lipids, Lipoproteins and Membranes, 5th edn. Amsterdam: Elsevier.
- Yeagle PL (2005) The Structure of Biological Membranes, 2nd edn.Boca Raton, FL: CRC Press.
Published date : 10 May 2014 03:16PM




More Articles


