

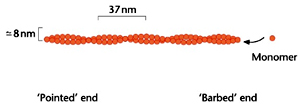
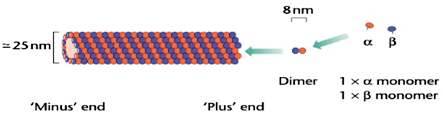
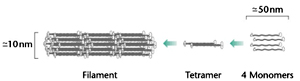
Microfilaments, the smallest, are about 8 nm in diameter. The largest fibres are microtubules, with a diameter of about 25 nm, which are so called because they clearly possess a hollow core in electron micrographs. Intermediate filaments have a diameter between that of microfilaments and microtubules, about 10 nm.
 These cytoskeletal elements are localized to different cellular parts according to their functions. Microfilaments are localized towards the periphery of the cell close to plasma membrane. They aid in bringing out rapid changes of the cell shape like in pseudopodia and ruffles. They are involved in cell crawling and motility. The hyaline (glass-like) appearances of Pseudopodia are often so rich in microfilaments that other, granular, components of the cytoplasm are excluded.
These cytoskeletal elements are localized to different cellular parts according to their functions. Microfilaments are localized towards the periphery of the cell close to plasma membrane. They aid in bringing out rapid changes of the cell shape like in pseudopodia and ruffles. They are involved in cell crawling and motility. The hyaline (glass-like) appearances of Pseudopodia are often so rich in microfilaments that other, granular, components of the cytoplasm are excluded.Microtubules, on the other hand, radiate from a point in the centre of the cell, near the nucleus, and only occasionally extend as far as the edge of the cell. The mitotic spindle that is responsible for dividing the duplicated chromosomes is largely composed of microtubules, suggesting that they have a role in moving and redistributing components within the cell.
 Most intermediate filaments are long, and form a meshwork spanning the whole width of cells formation of nuclear lamina. They provide strength to the cells in order to resist the wear and tear due to externally applied forces and are thus most prevalent in static cells, such as skin and heart muscle which experience continuous pressure.
Most intermediate filaments are long, and form a meshwork spanning the whole width of cells formation of nuclear lamina. They provide strength to the cells in order to resist the wear and tear due to externally applied forces and are thus most prevalent in static cells, such as skin and heart muscle which experience continuous pressure.All three cytoskeletal structures are assembled from small subunits called monomers which are polymerized to form long linear chains. The individual properties of these elements are derived from differences in the monomers and the way in which they are assembled.
Microfilaments and actin
Microfilaments are constructed from a single type of subunit, named actin. In its monomeric form, actin is usually referred to as G-actin (for globular actin). Each protein unit is about 375 amino acids long and 42 kDa in size, and contains either an ATP or ADP. Polymerization of these monomers occurs in head-to-tail fashion in an ATP dependent manner under high concentrations of K+ and Mg2+ to yield F-actin. F-actin is a tightly packed, simple helix containing just fewer than two subunits per turn and is polar. Since the monomers are all aligned in the same direction, the ends of the filament are chemically different, which is central to its behavior within cells.
Microtubules
Microtubules, on the other hand, are built from dimers of two 50-kDa proteins,a- and b-tubulin, each about 450 amino acids and 50 kDa in size. Tubulins use the guanine nucleotides guanosine triphosphate (GTP) and guanosine diphosphate (GDP) Instead of ATP and ADP,. Polymerization of dimmers is end to end, giving a polar filament. However, microtubules are composed of 13 filaments (protofilaments) encircling the hollow core, giving them far greater rigidity than actin microfilaments.
The flagella of protozoa and the cilia of vertebrates use a specialized assembly of tubulin and associated proteins to produce bending, whip-like movements. They exhibit a characteristic ‘9+2’ structure. An array of nine double-width microtubules, each formed like a figure of eight, is arranged radially around a core of two single microtubules.
Intermediate filaments
Intermediate filaments are very stable structures but are not homogeneous in composition and neither bind to nucleotides, nor use motors. They are made from a large family of related proteins like keratin family and neurofilaments ranging in size from 40 kDa to more than 100 kDa. Keratins make up tissues such as hair and nails, the family also includes a nerve-specific group called the neurofilaments, as well as desmin and vimentin. These proteins is found in a subset of differentiated cells in higher eukaryotes and are mostly absent in lower eukaryotes. Monomers assemble to form a rope-like structure, with several monomers intertwined together to make a structure with considerable tensile strength. Lamins constituting the nuclear lamina during interphase, are unique in attaining the form of latticework sheet after polymerization. Depolymerization of lamins leads to disppearance of nuclear envelope during Mitosis.
Actin microfilaments and microtubules are constantly being remodelled and redistributed within the cell. This continual change is fundamentally important to the working of the cytoskeleton. Nucleation is the key to new filament formation. Actin monomers or tubulin dimers are added to existing filaments to form the smallest functional filaments called ‘seeds’ or ‘nuclei’ which contain three monomers in the case of actin, or a ring of 13 tubulin dimers. Formation of new nuclei is a biphasic involving slow process initially, termed as lag phase followed by extension of the newly formed filaments termed as the rapid phase.
Mammalian cells in interphase contain only one microtubule organizing centre (MTOC), a structure that includes two centrioles fixed at right angles to each other. The centriole, like cilia and flagella, is made of nine fused ‘double’ microtubules forming a small cylinder. Surrounding the centrioles is a cloud of amorphous pericentriolar material. This entire structure is the centrosome. Higher land plants and some animal cells do not possess centrioles and so their MTOCs may be diffuse. Microtubules are initiated in the pericentriolar material (not from the centrioles) from a protein complex containing a third member of the tubulin family, g-tubulin. When the cell undergoes mitosis and starts to divide, a second centrosome must be constructed to provide one for each daughter cell. The new centrosome is not assembled independently; instead the original centrosome is somehow divided into two new daughters, which move apart towards the dividing halves of the cell. The mitotic spindle, the microtubule structure that actually separates the chromosomes, is then formed between the two new centrosomes.
Actin microfilaments, unlike microtubules, are nucleated by a mechanism that remains unknown despite decades of investigation.
Polarity and treadmilling
Polymerization of both actin and tubulin subunits are unidirectional in being head-to-tail or end to end and hence are said to exhibit polarity. Actin filaments have dissimilar ends one end is said to be 'barbed’ and the other end is ‘pointed’. For instance, the S1 fragment of myosin II binds to actin filaments at a constant angle where the myosin fragments are oriented like an arrow, with a point at one end and a barbed tail at the other. The end whose S1 fragments point outwards is therefore called the ‘barbed’ end, as opposed to the ‘pointed’ end in which the S1 points back along the filament. Polymerization is favoured at the barbed end as the actin filaments near the plasma membrane are all oriented with their barbed ends pointing outwards. This means that actin monomers usually appear to be added between the membrane and the end of the filaments.
Similarly ‘plus’ or growing ends of microtubules having points outwards from the MTOC, has a higher affinity for free tubulin than the ‘minus’ end. Tubulin dimers will tend to dissociate from the minus ends of all filaments, while at the same time adding to their plus ends. This simultaneous assembly of one end of a filament while disassembling the other has been called ‘treadmilling’. Energy is required for treadmilling in the form of a nucleotide triphosphate, ATP for actin and GTP for tubulin. Following polymerization, the ATP or GTP molecules are hydrolyzed to ADP or GDP, which drives a change in the binding affinity between subunits.
In cells microtubules they show another dynamic behaviour, called dynamic instability where the GTP bound to microtubules when gets hydrolyzed, depolymerization and shortening of the microtubule occurs at a catastrophic rate.
Severing
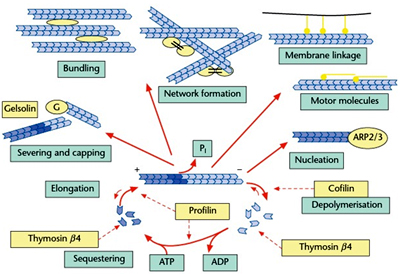
 Actin does not show dynamic instability. Instead, actin is disassembled by a family of severing proteins Actin-depolymerizing factor and its relatives, cofilin and actophorin are particularly associated with the dismantling of assembled actin microfilaments. Gelsolin is associated with both disassembly of existing actin filaments and nucleation of fresh ones.Actin binding proteins like villin align the actin filaments in the same direction (isotropically) into bundles to form long filaments in rigid fingers like structures called microvilli in epithelial cells increasing the surface area of absorption.
Actin does not show dynamic instability. Instead, actin is disassembled by a family of severing proteins Actin-depolymerizing factor and its relatives, cofilin and actophorin are particularly associated with the dismantling of assembled actin microfilaments. Gelsolin is associated with both disassembly of existing actin filaments and nucleation of fresh ones.Actin binding proteins like villin align the actin filaments in the same direction (isotropically) into bundles to form long filaments in rigid fingers like structures called microvilli in epithelial cells increasing the surface area of absorption.Filamin acts as a crosslinking protein that joins the actin fibers at an angle bringing more elasticity in lamellipodia (the active, ruffling, leading edge) of crawling cells.
Connections between the cytoskeleton and membranes
Actin cytoskeleton is connected to the plasma membrane with the aid of a number of proteins. The best characterized system is in red blood cells (erythrocytes), which require large surface area for efficient transfer of gases yet remain flexible enough to traverse through different capillaries. F-actin plays a key role in achieving the characteristic biconcave shape, which I turn is attached to a large complexes of several proteins. Two proteins, band III and glycophorin, are integral membrane-spanning proteins, and join the complexes to the membrane itself. Actin is bound by a large protein called spectrin, which is in turn connected to band III through ankyrin and to glycophorin through protein 4.1. Consequently, a compact crosslinked array of protein connecting multiple actin filaments to the whole area of the membrane is formed.
Motors: Proteins that Move Along Actin Filaments and Microtubules
Myosin II is a motor protein that acts as a part of highly ordered ‘thick filaments’ interdigitating the ‘thin filaments’ utilizing the energy from ATP to pull against actin filaments. Although most clearly visible in muscle, myosin II is present in essentially all eukaryotic cells.
Microtubules have their own individual motors, Kinesin and Dynein that play a major role in carrying vesicles and organelles along microtubules. Kinesin is a fairly large tetramer comprising two 120-kDa and two 62-kDa peptides. Whereas Dynein, an enormous molecule which contains two heavy chains of more than 470 kDa and several smaller subunits.
Extracellular signals like growth factors, chemotactic factors, extracellular matrix directly regulate the cytoskeleton’s form and movement. For instance in concentration of Calcium ions, an ubiquitous intracellular messenger plays a key role in muscle contraction. When the muscle is stimulated by nerve impulses, intracellular calcium channels open and the concentration rapidly rises bringing a change in the actin-binding proteins troponin and tropomyosin. Calcium binds directly to a subunit of troponin, and sufficient concentrations prevents it from inhibiting myosin II activity. Thus the rise in intracellular Ca2+ concentration activates the myosin II motor and allows the muscle to contract.
Chemotaxis and actin polymerization
Chemotaxis is the ability of different cells ranging from mammalian white blood cells to amoebae to move in the direction of attractive chemicals, chemo attractants. Growing nerves elongate towards peptides secreted by their targets, macrophages and neutrophils locate sites of infection by moving towards immune factors and bacterial metabolites, and location of the prey by amoebae constitutes a few examples of chemotaxis. Most chemotaxis is mediated by serpentine receptors and heterotrimeric G-proteins, the larger relatives of small GTP-binding proteins. When the receptors bind an appropriate ligand, the actin cytoskeleton is rapidly reorganized. In resting cells around 30–40% of actin monomers are assembled into filaments, but this amount can double within a few seconds of a chemoattractant being added.
References
- Murakami et.al, (2010) Structural basis for actin assembly, activation of ATP hydrolysis and delayed phosphate release. Cell 143; 275-287.
- Tondeleir D et.al, (2009) Actin isoform expression patterns during mammalian development and in pathology; insights from mouse models. Cell Motility and Cytoskeleton 66; 798-815.
- Pollard TD and Borisy GG (2009) Actin, a central player in cell shape and movement. Science 326; 1208-1212.
- Desai and Mitchinson TJ (1998) Microtubule polymerization and dynamics. Annual Review of Cell and Development Biology 13:197-231.
- Schmidt A and Hall MN (1998) Signaling to the actin cytoskeleton. Annual Review of Cell and Development Biology 14:305-338.
| Comparision of cytoskeletal elements | ||||
| S.No | Property | Microfilaments | Microtubules | Intermediate filaments |
| 1 | Size | 8nm | 25nm | 10nm |
| 2 | Shape | Filamentous, cable like | Filamentous, Tube like (Hollow inside) | Long meshlike |
| 3 | Stability | Dynamic (constantly remodeled) | Dynamic (constantly remodeled) | Stable |
| 4 | Cellular localization | Peripheral, near the plasma membrane | Central, near to the nucleus | Spread all over the cell |
| 5 | Function involved | Cell motility, Cell absorption, Muscle contraction, Chemotaxis. | Cell division, Intracellular transport and locomotory organs. | Cell structure and strengthening. |
| 6 | Type of subumits | Globular monomer, | Dumbbell shaped Heterodimer , | Varying (Keratin and neurofilament family) |
| 7 | Polymerisation | Head to tail fashion (polar) | End to end (polar) | side by side into rope-like structures |
| 8 | Smallest functional unit | Made up of three | Made up of 13 protofilaments | Non homogenous (varying) |
| 9 | Affinity to nucleotides | Bind to ATP/ADP | Bind to GTP/GDP | Do not bind to nucleotides. |
| 10 | Motor proteins | Myosin | Kinesin and Dyenin | Do not bind to motors |
| 11 | Treadmilling | Absent | shown | Absent |
| 12 | Dynamic Instability | Absent | shown | Absent |
| 13 | Severing | Shown | absent | Absent |
More Articles

