
Sakshi Education

Fatty acids have four major physiological roles:
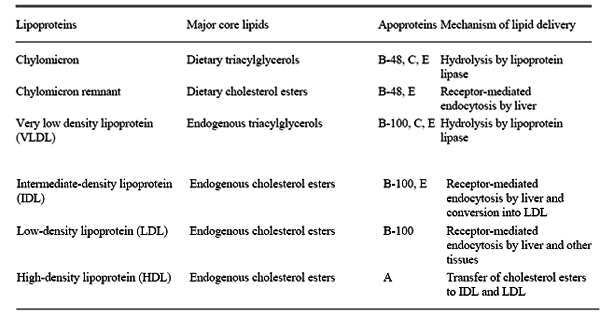
Lipases secreted by the pancreas convert triacyl glycerols into fatty acids and monoacylglycerol for absorption into the intestine.
Free fatty acids and monoacylglycerols are absorbed by intestinal epithelial cells. Triacylglycerols are resynthesised and packaged with other lipids and apoprotein B-48 to form chylomicrons, which are then released into the lymph system and then into the blood. These particles bind to membrane-bound lipoprotein lipases, primarily at adipose tissue and muscle, where the triacylglycerols are once again degraded into free fatty acids and monoacylglycerols for transport into the tissue. The triacylglycerols are then resynthesized inside the cell and stored. In the muscle, they can be oxidized to provide energy.
Utilization of Lipids: The utilization of Fatty acids as fuel requires three stages of processing - 
Fatty acids are activated before they enter into the mitochondrial matrix. This activation reaction takes place on the outer mitochondrial membrane, where it catalysed by acyl CoA synthetase (also called fatty acid thiokinase).
Fatty acids are activated on the outer mitochondrial membrane, whereas they are oxidized in the mitochondrial matrix. A special transport mechanism is needed to carry long-chain fatty acids are transported across inner mitochondrial membrane. Activated long chain fatty acids are transported across the membrane by conjugating them to carnitine, a zwitterionic alcohol. The acyl group is transferred from the sulfur atom of CoA to the hydroxyl group of carnitine to form acyl carnitine. the membrane by conjugating them (acyl carnitine transferase I)

The entry of acyl carnitine into the mitochondrial matrix is mediated by a translocase. Carnitine returns to the cytosolic side of the inner mitohondrial membrane in exchange for acyl carnitine.
Oxidation of Fatty acids:


The oxidation of unsaturated fatty acids present some difficulties, yet many such fatty acids are available in the diet. Most of the reactions are the same as those for saturated fatty acids. In fact, only two additional enzymes – an isomerase and a reductase – are needed to degrade a wide range of unsaturated fatty acids. Fatty acids having an odd number of carbon atoms are minor species. They are oxidized in the same way as fatty acids having an even number, except that propionyl CoA and Acetyl CoA, rather than two molecules of acetyl CoA, are produced in the final round of degradation. The activated three carbon unit in propionyl CoA enters the citric acid cycle after it has been converted into succinyl CoA that requires vitamin B12 as its coenzyme.
Ketone Bodies: Acetoacetate, d-3-hydroxybutyrate and acetone are often referred to as ketone bodies. Abnormally high levels of ketone bodies are present in the blood of untreated diabetics.

The major site of production of acetoacetate and 3-hydroxybutyrate is the liver. These substances diffuse from the liver mitochondria into the blood and are transported to peripheral tissues. Acetoacetate and 3-hydroxybutyrate are normal fuels of respiration and are quantitatively important as sources of energy. Indeed, heart muscle and the renal cortex use acetate in preference to glucose. In contrast, glucose is the major fuel for the brain and red blood cells in well-nourished people on a balanced diet. However, the brain adapts to the utilization of acetoacetate during starvation and diabetes. In prolonged starvation, 75% of the fuel needs of the brain are met by ketone bodies.
In case of insulin dependent diabetes mellitus production of large amounts of ketone bodies occurs in the liver, which are moderately strong acids. The result is severe acidosis (Diabetic ketosis). The decrease in pH impairs tissue function, most importantly in the central nervous system.
Fatty acid synthesis: 
The source of Acetyl CoA:

Hormones:
Archidonate, a 20:4 fatty acid derived from linoleate, is the major precursor of several classes of signal molecules: prostaglandins, prostacyclins, thromboxanes, and leukotrienes.

Prostaglandins, prostacyclin, thromboxanes, and leukotrienes (first found in leukocytes, contain three conjugated double bonds - hence the name) are called eicosanoids (from the Greek eikosi, “twenty”) because they contain 20 carbon atoms.
Prostaglandins stimulate inflammation, regulate blood flow to particular organs, control ion transport across membranes, modulate synaptic transmission, and induce sleep. Drugs such as aspirin and ibuprofen block the channel and prevent prostaglandin syntheis by inhibiting cyclooxygenase activity of the synthase.
- Fatty acids are building blocks of phospholipids and glycolipids. These amphipathic molecules are important component of biological membranes
- Many proteins are modified by the covalent attachment of fatty acids, which targets them to membrane locations.
- Fattyacids are fuel molecules. They are stored as triacylglycerols.
- Fatty acid derivatives serve as hormones and intracellular messengers
Lipases secreted by the pancreas convert triacyl glycerols into fatty acids and monoacylglycerol for absorption into the intestine.
Free fatty acids and monoacylglycerols are absorbed by intestinal epithelial cells. Triacylglycerols are resynthesised and packaged with other lipids and apoprotein B-48 to form chylomicrons, which are then released into the lymph system and then into the blood. These particles bind to membrane-bound lipoprotein lipases, primarily at adipose tissue and muscle, where the triacylglycerols are once again degraded into free fatty acids and monoacylglycerols for transport into the tissue. The triacylglycerols are then resynthesized inside the cell and stored. In the muscle, they can be oxidized to provide energy.
Utilization of Lipids: The utilization of Fatty acids as fuel requires three stages of processing -
- Mobilization of fatty acids from adipose to the energy requiring tissues.
- The fatty acids must be activated and transported into mitochondria for degradation.
- Fatty acids are broken down in a step-by-step fashion into acetyl coA, which is then processed in the citric acid cycle.
- Mobilization of Fatty acids:

Triacyl glycerols in adipose tissue are converted into free fatty acids and glycerol (a process known as lipolysis) for release into the blood stream in response to hormonal signals (epinephrine, norepinephrin, glucagons and adrenocorticotrophic haormone). In contrast, insulin inhibit lipolysis. The released fatty acids are not soluble in blood plasma, and so, on release, serum albumin binds free fatty acids and serves as a carrier.
Glycerol formed by lipolysis is absorbed by the liver and phosphoylated, oxidized to dihydroxyacetone phosphate, and then isomerised to glyceraldehydes-3-phosphate. So, glycerol can be converted into pyruvate or glucose in the liver and the reverse process also occur. Thus, glycerol and glycolytic intermediates are readily interconvertable.

- Fatty acid activation and Transportation into the mitochondria:
Fatty acids are activated before they enter into the mitochondrial matrix. This activation reaction takes place on the outer mitochondrial membrane, where it catalysed by acyl CoA synthetase (also called fatty acid thiokinase).
Fatty acids are activated on the outer mitochondrial membrane, whereas they are oxidized in the mitochondrial matrix. A special transport mechanism is needed to carry long-chain fatty acids are transported across inner mitochondrial membrane. Activated long chain fatty acids are transported across the membrane by conjugating them to carnitine, a zwitterionic alcohol. The acyl group is transferred from the sulfur atom of CoA to the hydroxyl group of carnitine to form acyl carnitine. the membrane by conjugating them (acyl carnitine transferase I)
The entry of acyl carnitine into the mitochondrial matrix is mediated by a translocase. Carnitine returns to the cytosolic side of the inner mitohondrial membrane in exchange for acyl carnitine.
Oxidation of Fatty acids:
The oxidation of unsaturated fatty acids present some difficulties, yet many such fatty acids are available in the diet. Most of the reactions are the same as those for saturated fatty acids. In fact, only two additional enzymes – an isomerase and a reductase – are needed to degrade a wide range of unsaturated fatty acids. Fatty acids having an odd number of carbon atoms are minor species. They are oxidized in the same way as fatty acids having an even number, except that propionyl CoA and Acetyl CoA, rather than two molecules of acetyl CoA, are produced in the final round of degradation. The activated three carbon unit in propionyl CoA enters the citric acid cycle after it has been converted into succinyl CoA that requires vitamin B12 as its coenzyme.
Ketone Bodies: Acetoacetate, d-3-hydroxybutyrate and acetone are often referred to as ketone bodies. Abnormally high levels of ketone bodies are present in the blood of untreated diabetics.
The major site of production of acetoacetate and 3-hydroxybutyrate is the liver. These substances diffuse from the liver mitochondria into the blood and are transported to peripheral tissues. Acetoacetate and 3-hydroxybutyrate are normal fuels of respiration and are quantitatively important as sources of energy. Indeed, heart muscle and the renal cortex use acetate in preference to glucose. In contrast, glucose is the major fuel for the brain and red blood cells in well-nourished people on a balanced diet. However, the brain adapts to the utilization of acetoacetate during starvation and diabetes. In prolonged starvation, 75% of the fuel needs of the brain are met by ketone bodies.
In case of insulin dependent diabetes mellitus production of large amounts of ketone bodies occurs in the liver, which are moderately strong acids. The result is severe acidosis (Diabetic ketosis). The decrease in pH impairs tissue function, most importantly in the central nervous system.
Fatty acid synthesis:
- Synthesis takes place in the cytosol, in contrast with degradation, which takes place primarily in the mitochondrial matrix
- Intermediates in fatty acid synthesis are covalently linked to the sulfhydryl groups of an acyl carrier protein (ACP), whereas intermediates in fatty acid breakdown are covalently attached to the sulfhydryl group of coenzyme A
- The enzymes of fatty acid synthesis in higher organisms are joined in a single polypeptide chain called fatty acid synthase. In contrast, the degradative enzymes donot seem to be associated.
- The growing fatty acid chain is elongated by the sequential addition of two-carbon units derived from acetyl CoA. The activated donor of two carbon units in the elongation step in malonyl ACP. The elongation reaction is driven by the release of C02.
- The reductant in fatty acid syntheis is NADPH, whereas the oxidants in fatty acid degradation are NAD+ and FAD.
- Elongation by the fatty acid synthase complex stops on formation of pamitate (C16). Further elongation and the insertion of double bonds are carried out by other enzyme systems.
The source of Acetyl CoA:
Hormones:
Archidonate, a 20:4 fatty acid derived from linoleate, is the major precursor of several classes of signal molecules: prostaglandins, prostacyclins, thromboxanes, and leukotrienes.
Prostaglandins, prostacyclin, thromboxanes, and leukotrienes (first found in leukocytes, contain three conjugated double bonds - hence the name) are called eicosanoids (from the Greek eikosi, “twenty”) because they contain 20 carbon atoms.
Prostaglandins stimulate inflammation, regulate blood flow to particular organs, control ion transport across membranes, modulate synaptic transmission, and induce sleep. Drugs such as aspirin and ibuprofen block the channel and prevent prostaglandin syntheis by inhibiting cyclooxygenase activity of the synthase.
Published date : 23 May 2014 02:08PM




More Articles


Most Read
